Most of our prior work focused on the Ubl protein SUMO as it is essential in budding yeast and its conjugation to substrates requires an enzyme cascade that involves fewer factors in comparison to the ubiquitin conjugation cascade thus facilitating both structural and functional studies. Our more recent efforts now encompass both SUMO and ubiquitin in studies that address their activation, conjugation and recognition.
E1 activation
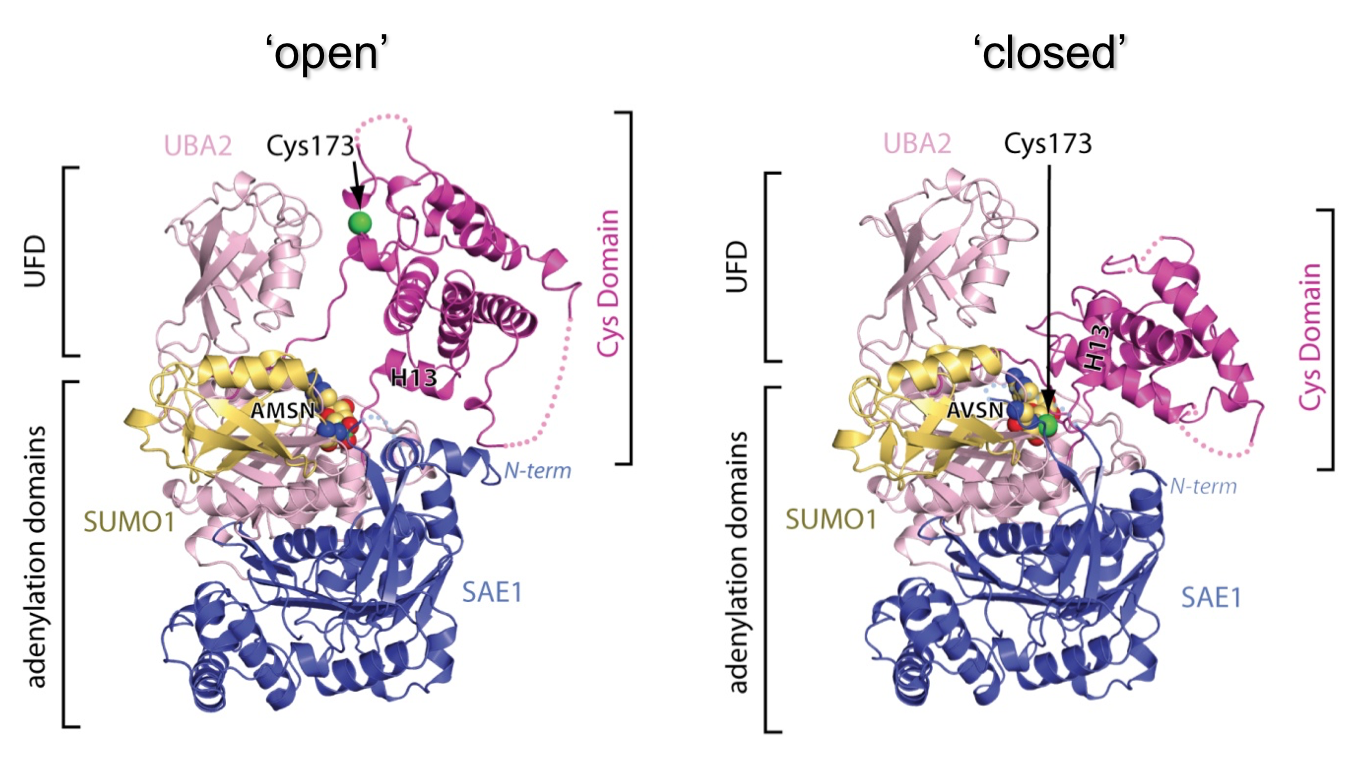
E1 enzymes facilitate conjugation of ubiquitin and ubiquitin-like proteins through adenylation, thioester transfer within E1, and thioester transfer from E1 to E2 conjugating proteins. Early work determined structures of the SUMO E1 in complex with SUMO and ATP and elements required for E2 recruitment, however the E1 active site cysteine was positioned to far from the SUMO C-terminus to explain the second reaction, thioester bond formation, suggesting that additional changes within the adenylation site were required to promote adenylation and subsequent thioester transfer. Subsequent work revealed the structural basis for requisite conformational changes through application of chemical probes that mimicked unstable intermediates in the pathway, showing that the E1 active site is uniquely remodeled to promote adenylation and thioester bond formation. We also determined the structural basis for ubiquitin E1-E2 thioester bond formation.
Substrate recognition by the E2
The structural and biochemical basis for E2-dependent protein conjugation was uncovered by analysis of a complex between human Ubc9 and RanGAP1. These studies revealed structural determinants for recognition of consensus SUMO modification sequences found within SUMO conjugated proteins. Reconstituting SUMO conjugation in vitro revealed distinct motifs in the E2 required for substrate binding and SUMO modification of various substrates. These studies were followed by an in vivo screen in yeast that uncovered at least three critical E2 residues that were important for function in vivo and for conjugation in vitro. Interestingly, kinetic analysis of the mutant and wild-type SUMO E2 revealed that the E2 can activate the substrate for conjugation, a mechanism that is likely application to E2-mediated conjugation in other Ub/Ubl pathways.
E3 ligases
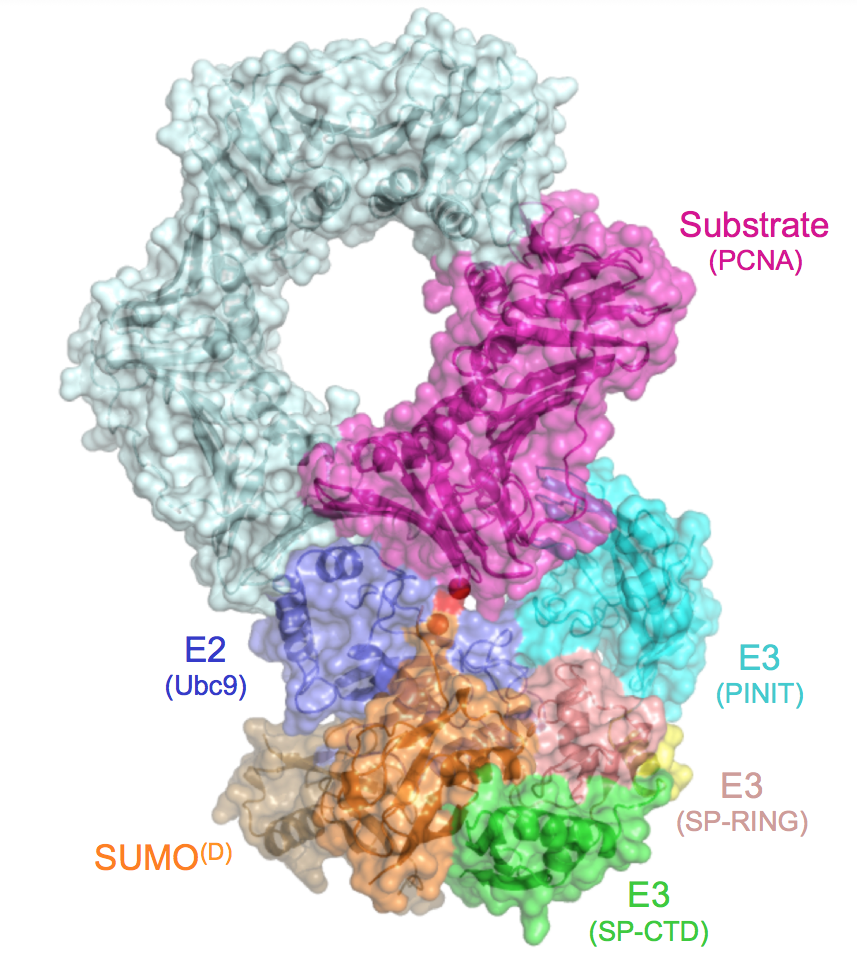
Unique combinations of E2s and E3s ensure specificity and enhance conjugation in the ubiquitin pathway. The SUMO pathway includes only one known E2 and just a few E3s. We employ structural and biochemical tools to uncover the basis for substrate specificity for non-RING and RING E3s that differ in their ability to selectively target substrates for modification. One early study revealed the kinetic basis for E3 ligase domains within the Nup358/RanBP2 nucleoporin, including structural characterization of a complex between human Nup358/RanBP2, the SUMO E2 Ubc9, and SUMO-1 conjugated RanGAP1. This study revealed that the E3 organized the charged E2-thioester to promote conjugation, a feature shared with many other ubiquitin and ubiqutin-like E3 ligases. Subsequent studies revealed a structural basis for the ZNF451 SUMO E3 ligase, and the structural basis for substrate recognition in an activated RING-type SUMO E3 ligase.
Recognition of Ub/Ubl-modified proteins
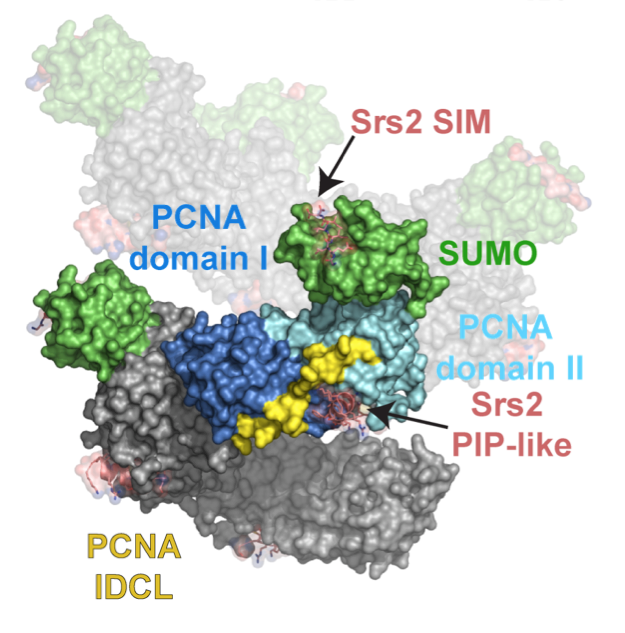
Ubiquitin and ubiquitin-like proteins are conjugated to other proteins to generate signals. In many instances, the Ub/Ubl-modified protein is specifically recognized by downstream factors. We focused on recognition of SUMO-modified PCNA, the trimeric ring that encircles DNA to facilitate replication and DNA repair. In budding yeast, SUMO-PCNA signals for recruitment of the anti-recombinogenic helicase Srs2 and we were able to determine that its C-terminal domain harbored tandem motifs, one for SUMO binding and one for PCNA binding, that were required for specific recognition of SUMO-PCNA in vitro and in vivo. Ongoing studies are aimed at addressed how other receptors specifically ’see’ Ubl conjugated substrates to elicit downstream signalling events.
Proteases in Ub/Ubl pathways
Equally important in Ub/Ubl pathways is the process of deconjugation, to remove the Ubl signal after its completed its function. Deconjugation is catalyzed by unique proteases that recognize the respective Ubl protein and/or substrate. We characterized several of SUMO proteases through the structure determination, biochemical, and genetic analysis of the complexes in yeast between S. cerevisiae and human. In addition, we also characterzed a ’ulp’ type protease that is able to deconjugate Nedd8 rather than SUMO and a viral protease that removes lysine 48-linked ubiquitin chains two ubiquitins at a time.